|
魚類腸道正常菌群研究進展 宋增福 1,吳天星2 (1.上海水產大學生命學院,上海 200090;2.浙江大學化學系,浙江 杭州 310027) 關鍵詞:魚;腸道;正常菌群 作者簡介:宋增福(1971-),男,博士 研究方向:水產微生態與疾病防治;E-mail:zfsong@shfu.edu.cn. 通訊作者:吳天星(1963-),男,教授,博士生導師 研究方向:動物營養學與飼料科學;E-mail:wutx@tianbang.com
魚類腸道正常菌群是腸道的正常組成部分;是腸道微生物與宿主以及所處的水生環境形成的相互依賴、相互制約的微生態系;對營養物質的消化吸收、免疫反應以及器官的發育等方面具有其他因素不可替代的作用,并且影響到魚類的生長、發育、生理和病理。筆者擬就魚類腸道菌群的形成、結構與數量、生理功能以及影響菌群結構的因素、與益生菌的關系等方面加以綜述。
1 魚腸道正常菌群的形成
研究表明細菌最初的定植過程在幼魚發育階段是非常復雜的,通常受到多種因素的影響,但主要是決定于魚卵表面、活的餌料和幼魚飼養水體中的細菌[122]。處于孵化階段的幼魚具有一個發育不完全的消化道,其內是無菌的。處于孵化過程中的幼魚主要依靠卵黃來供給營養物質,當其從卵中孵化出來,一旦接觸到周圍的水生環境和活餌料,多種細菌就開始在腸道上皮定植[327]。Mroga等[6]研究發現腸道菌群的主要來源是所攝取的活飼料而不是養殖水體。另有結果表明,最先定植的細菌能調節上皮細胞的基因表達,從而使最先定植的細菌與宿主腸道環境相適應,并且可以阻止在這個生態系統中后來的細菌的定植。因此,最初的細菌定植與成年最終穩定的腸道菌群組成結構具有高度相關性。然而,也有的試驗結果表明腸道的菌群與魚的飼料和水體環境中的細菌并不相同[8]。
2 魚類腸道菌群的特性、組成及數量
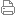
魚類腸道菌群細菌種類繁多,數量極大。有研究報道指出,淡水魚腸內細菌的數量基本為105~108[9],而海水魚腸內細菌的數量106~108[10]。腸道的優勢細菌為革蘭氏陰性菌,同時也存在革蘭氏陽性菌[11]。 由于魚類的生存生長環境與陸生動物不同,因此,在腸道微生態系中其細菌的某些生理生化特征也表現出特異性。Smith對魚的腸道的大腸桿菌進行研究時就發現,魚腸道的大腸桿菌可以液化明膠,不產生吲哚,而這些特性是從陸生動物腸道分離的大腸桿菌所不具備的。 不同種類的魚之間,由于所處的水體環境、食性等因素,其腸道細菌組成和結構也不盡相同的。研究表明淡水魚類腸道內專性厭氧菌以A、B型擬桿菌科等為主[11212],好氧和兼性厭氧細菌則以氣單胞菌屬、腸桿菌科等為主[9]。乳酸菌在陸生動物是常駐菌,而在魚類也是腸道菌群的組成部分。Ring等[13219]曾對乳酸菌進行了系統研究。王紅寧[20]對淡水養殖池中的鯉魚腸道的菌群結構研究發現,在鯉魚腸道中的需氧和兼行性厭氧菌的數量依次為:氣單胞菌、酵母菌、大腸桿菌、假單胞菌、葡萄球菌、需氧芽孢桿菌。氣單胞菌和酵母菌的數量更多。可以認為是腸道里的優勢需氧、兼性厭氧菌。厭氧菌的數量依次是:擬桿菌、乳酸桿菌、梭狀芽孢桿菌,其中擬桿菌數量最多,可以認為是腸道中的優勢厭氧菌。尹軍霞等[21]對淡水養殖池中的4種不同食性魚—烏鱧、鰱、鳊、鯽的腸道壁菌群進行了定性、定量分析,發現不論是好氧菌還是厭氧菌,同種魚前腸壁分布一般比中腸壁和后腸壁少;同一腸段相比,都是厭氧菌總數遠大于好氧菌總數,一般相差2~3個數量級;不同魚種之間,腸壁的好氧菌總數差別比厭氧菌總數差別大得多;厭氧菌中的乳酸球菌和雙歧桿菌具有一定的正相關性。4種魚腸道壁中的厭氧菌總數和雙歧桿菌分布的規律是:肉食性的烏鱧>雜食性和廣食性的鯽>食浮游植物為主的鰱>草食性的鳊,即魚類腸道壁中的厭氧菌總數和雙歧桿菌隨著從草食性向肉食性發展而逐漸增加。Rachel等[22]從淡水扁鯊和Oscars以及南方比目魚中分離到梭菌、革蘭氏陰性菌屬的梭菌、擬桿菌等細菌。因此,魚類腸道的菌群組成結構隨著魚種類、食性、生長的環境的不同而呈現出差異。
3 魚類腸道正常菌群的生理功能
腸道正常菌群在魚類的生長發育過程中擔當非常重要的作用,它既要參與營養物質的消化和吸收,同時又要擔當機體的防御功能,維護機體的健康。 3.1 營養功能 根據微生態的三流運轉理論,微生態系統中存在能源流動、物質交換和基因傳遞。動物、人類及植物的組織細胞與正常微生物之間以及正常微生物與正常微生物之間都存在著能源的交換。電鏡觀察發現腸上皮細胞表面的微絨毛與菌體細胞壁上的菌毛極為貼近,并有物質交換的跡象。 在魚類腸道微生態環境中,正常菌群的建立通常被認為是對動物發育不完全腸道酶系的有益補充,尤其是在幼魚發育階段。它能合成分泌一些天然食物中不含有而宿主生長發育所必需的維生素、酶、調節因子等,且能分解產生一些易被宿主吸收的物質供宿主吸收。同時,正常菌群及其某些新陳代謝產物能促進宿主對某些營養物質的吸收,改變宿主消化酶的活性,從而對宿主的新陳代謝起著不可缺少的影響作用。Sugita等[23]從羅非魚和鯰魚腸道里分離出具有產生維生素B12的細菌。早期對胭脂魚的研究表明,從腸道內分離的弧菌和腸道菌具有分泌蛋白酶、淀粉酶、幾丁質酶和卵磷脂酶的能力[24]。Das等[25]研究草魚消化酶時,為校正腸道細菌分泌的酶所引起的誤差,用抗菌素飼喂草魚,然后比較試驗組與對照組魚腸道狡甲基纖維素分解菌,發現腸道細菌的纖維素酶對草魚消化食物中的纖維素起了一定的促進作用。Sugita等[26]分析了香魚、日本鰻鱺、鯉、羅非魚、斑點叉尾鲴回等淡水魚類腸道菌群的產酶能力,認為腸道菌群在淡水魚的淀粉消化中占有重要的地位。潘康成等[27]利用2種有益地衣芽孢桿菌,飼喂鯉魚,對鯉魚的生長有明顯地促進作用和顯著提高鯉魚腸道消化酶的活性。Bachel等[22]分析了淡水扁鯊、Oscars和南方比目魚的腸道厭氧菌的產酶功能,發現這些腸道的厭氧菌可以產生酸性和堿性磷酸酶、四碳和八碳的酯酶、十四碳的脂肪酶、芳基化合物分解酶和糖苷酶等。Schrijiver[28]研究發現,當日糧中添加解蛋白弧菌時,解蛋白弧菌會提高表觀氮消化率(P<0.01)。由此可見,腸道菌群分泌的蛋白酶、淀粉酶等消化酶可以輔助魚類營養物質的消化,包括一些難以分解的纖維素等物質,從而促進營養物質的轉化和吸收;產生的維生素等營養物質可以彌補飼料中營養物質的不足。這就為外源益生菌的添加會促進魚的生長提供了科學依據。 3.2 防御功能 正常微生物群覆蓋在腸道黏膜的表面作為物理屏障可以拮抗致病菌的入侵;同時可以刺激機體產生非特異性和特異性免疫反應從而加強機體的保護作用。這種防御機制主要包括定植阻抗、病原菌抑制和免疫調節作用。 3.2.1 定植阻抗 在動物腸道中,正常菌群其厭氧菌占99%,它們是構成動物腸道屏障之一。定植阻抗是指腸道菌群具有排斥病原菌在腸道上皮細胞定植的作用,也可以形象地理解為腸道菌群就像柵欄一樣起到了阻攔病原菌的作用。動物模型試驗在這方面提供了有力的證據。20世紀60年代的小鼠沙門氏菌感染試驗最具有說服力。試驗結果表明,無菌鼠的沙門氏菌致死量只有普通鼠的十億分之一。由此可見,腸道菌群的屏障保護作用非常顯著。Bergh[29]指出,魚類的體表和消化道黏膜內定居著共生性細菌,他們形成菌叢,從而成為防止病原菌定居和增殖的屏障。 3.2.2 病原菌抑制 試驗證明,在從魚腸道里分離的部分細菌具有抑菌的作用,可以保護宿主免受病原菌的侵害。Sugita等[26]對從魚類的腸道菌群中分離出304株細菌進行抗菌的試驗研究發現,3.23%表現出抗菌的能力,尤其是嗜水單胞菌、假單胞菌、梭狀芽孢桿菌的幾個菌株可以抑制7~12個靶菌株。在此基礎上的抗菌效果顯示,抗菌活力隨魚的種類、采樣時間、部位而有差別,同時這些抗菌物質對腸道菌群的組成也有影響。Olsson[30]用鰻弧菌飼喂大菱鲆,發現腸道肉桿菌可以抑制鰻弧菌的生長。 3.2.3 免疫調節 魚類免疫反應是一種極為復雜的生理現象,受很多環境因素的影響。魚類在進行自身免疫保護時,主要行使非特異性(先天性)與特異性(獲得性)兩大免疫防御功能。 李莉[31]研究發現,隨著腸道內氣單胞菌(Aeromonas)百分含量的下降,腸擬桿菌(Enterobacteriaceae)百分含量的上升,草魚白細胞的吞噬百分比和吞噬指數明顯上升,這說明草魚腸道優勢菌群氣單胞菌、腸擬桿菌的變化可能與白細胞吞噬活性增強有關。Panigrahi[32發現,虹鱒魚飼喂乳酸菌(LactobacillusrhamnosusJCM)時,活的乳酸菌比熱致死的乳酸菌能夠更好地誘導巨噬細胞的吞噬活性和補體功能,并且使血漿免疫球蛋白水平呈現增加趨勢。因此,魚類腸道菌群的變化或者外來益生菌的添加均會引起機體的免疫反應,但是他們之間內在的關系尚不清楚,還有待于更多的試驗探討。
4 影響魚腸道菌群結構與組成的因素
動物微生態系統是由動物、微生物和環境彼此相互作用、相互依賴的統一整體,這個整體是由不同層次、不同環節相互聯結成的立體交叉網絡,這個網絡就是微生態系統的多維空間結構。作為魚的腸道微生物菌群,由于所處的宿主的腸道生長環境是和周圍的水體環境緊密相關,形成一個整體,其平衡更容易受到外界水體環境變化、餌料的變換等因素的影響,尤其是在幼苗時期。Cahill[33]曾就眾多的試驗結果總結認為,水環境中的細菌影響腸道微生物的菌群結構,同時腸道的菌群結構也影響水體環境的微生物結構,腸道里的微生物是那些和環境中的微生物相同的種類或者是那些在日糧中存在的并且能在腸道繁殖的微生物。Ring等[14]曾對腸道的乳酸菌進行了細致的研究,發現不飽和脂肪酸、營養競爭、三氧化二鉻、鹽度和應激等因素都會對腸道的乳酸菌產生影響。因此,與魚直接接觸的水體環境對魚腸道微生物菌群的影響要遠遠大于陸生動物和人所處的外部環境對腸道微生物菌群的作用。 魚類是變溫動物,消化道的微生物也會隨體溫的變化而發生變化。曾勇等[34]研究了草魚腸道中氣單胞菌的季節變化。在冬季停食階段,前腸壁的氣單胞菌的占總菌數比例明顯升高。Janina等[35]研究發現,在3月未檢測到比目魚腸道的菌群,而在夏天檢測到細菌數為14.5×106。 飼料的組成能改善魚消化道微生物菌群的結構。曹志華等[36]的研究發現,飼喂鯉魚含有尿素的飼料,對照組腸道中的優勢菌為氣單胞菌、芽孢桿菌、葡萄球菌和微球菌,可利用尿素的細菌占細菌總數的55%。試驗組的優勢菌為芽孢桿菌,腸桿菌和葡萄球菌,可利用尿素的細菌占85%。陳孝煊[37]的研究發現,飽食狀態下草魚的腸道菌的平均為8.7×107cfu/g,空腸狀態下為7.29×106cfu/g,差異顯著。 在正常生理條件下,腸道正常的菌群會發生變化;在病理條件下,腸道菌群的變化往往是病因或結果。在發病的時候,條件致病菌大量繁殖,會導致腸道菌群的平衡失調,進而導致整個病情的惡化。蔣長苗等[38]研究了草魚腸道正常菌群與腸炎病原菌間的關系,發現發生腸炎的草魚,點狀產氣單胞菌、大腸桿菌、腸球菌顯著增加,而乳酸桿菌、雙歧桿菌數量減少,厭氧菌與需氧菌比值顯著下降。從微生態學的角度認為,以厭氧菌顯著下降為特征的菌群失調是草魚腸炎的發病機制之一。
5 研究魚類腸道菌群試驗動物模型和方法
對魚腸道正常菌群的研究常用和有效的方法是傳統的細菌純培養的方法,但是該方法常規培養的腸道細菌的種類有限,而且此方法工作量大,分析數據不夠準確。隨著分子生物學的迅速發展,將為魚腸道的菌群分析提供了有力的技術支撐。 5.1 新型試驗動物模型的建立為研究魚的腸道的細菌提供了有利的工具 無菌動物在腸道的細菌研究中發揮著重要的作用,尤其是在陸生動物研究方面。目前大多數的有關腸道菌群的知識是通過無菌動物的研究獲得的。但是在魚類腸道的研究方面,由于沒有無菌魚,因此在研究時就顯得有些力不從心,所以很多的相關知識是從陸生無菌動物的研究中借鑒而來的。無菌魚的出現使得這種狀況得以改變。 5.2 分子生物學的方法為研究魚的腸道正常菌群開辟了新的途徑 用分子生物學方法研究人和動物的腸道菌群早已得到應用。而在魚類的腸道菌群研究方面,這些方法應用比較少,如核酸探針、原位雜交,變性梯度電泳等。但是這將為魚類腸道正常菌群的研究開辟新的途徑。
6 益生菌對魚類腸道菌群的調控
魚類的腸道微生物區系在魚的健康方面扮演非常重要的角色。腸道的微生物區系的動態平衡是動物保持個體健康和發揮正常生產性能所必需的條件,調控魚類消化道微生物區系對提高魚類的生產性能和飼料利用率,減少病原菌在腸道內的定植具有重要的意義。益生菌調控腸道微生物區系的作用已越來越受到人們的重視。王紅寧[20]研究了微生態添加劑飼喂鯉魚腸道內的7種菌群(氣單胞菌、大腸桿菌、需氧芽孢桿菌、酵母菌、乳酸桿菌、雙歧桿菌和擬桿菌)并對這些菌群進行了定量和定性分析,結果表明試驗組的大腸桿菌數顯著<對照組,試驗組的氣單胞菌、需氧芽孢桿菌、酵母菌、雙歧桿菌和擬桿菌有所增多,但是與對照組無顯著差異,而乳酸桿菌略有減少。可見微生物添加劑有顯著抑制鯉魚腸道大腸桿菌的作用,并使腸道內的微生物數量通過其他微生物(有益微生物的添加)而達到新的平衡狀態。 魚類腸道菌群的平衡是魚腸道發揮正常生理作用的外在基礎,因此,積極研究和探討魚類腸道的微生態并加以調控,使之建立穩定平衡的菌群體系,對維護魚類的健康,提高魚的產量,生產安全的水產品,具有積極的理論和實踐意義。益生菌作為調整腸道菌群平衡的活菌制劑,必將會在水產業中發揮積極的作用。
參考文獻: [1]
OlafsenJA.Igestionofbacteriabycod(GadusmorhuaL.)lavae[G]//ThePropagationofCodGadusmorhuaL.Instituteofmarineresearch,FlodevigenBiologicalStation,1984:6272643. [2]
HansenGH,OlafsenJA.Bacterialcolonizationofcod(GadusmorhuaL.)andhalibut(Hippoglossushippo2glossus)eggsinmarineaquaculture[J].Appliedandenvironmentalmicrobiology,1989(55):143521446. [3]CampbellAC,BuswellJA.TheintestinalmicrofloraoffarmedDoversoleSoleasoleaatdifferentstagesoffishdevelopment[J].JApplBacteriol,1983(55):2152223. [4]MurogaK,HigashiM,KeitokuH.Theisolationofin2testinalmicrofloraoffarmedredseabreamPagrusmajorandblackseabreamAcanthopagrusschlegeliatlarvalandjuvenilestages[J].Aquaculture,1987(65):79288. [5]HansenGH,StrmE,OlafsenJA.EffectofdifferentholdingregimesontheintestinalmicrofloraofherringClupeaharenguslarvae[J].ApplEnvironMicrobiol,1992(58):4612470. [6]MunroPD,BarbourA,BirkbeckTH.Comparisonofthegutbacterialfloraofstart2feedinglarvalturbotrearedunderdifferentconditions[J].JApplBacteriol,1994(77):5602566. [7]RingE,BirkbeckTH,MunroPD,etal.TheeffectofearlyexposuretoVibriopelagiusontheaerobicbacterialfloraofturbotScophthalmusmaximusL.larvae[J]J ApplBacteriol,1996(81):2072211. [8]NicolasJL,RobicE,AnsqerD.Bacteialfloraassociat2edwithatropicchainconsistingofmicroalgaerotifersandturbotlarvae:influencesofbacteriaonlarvalsurvival[J].Aquaculture,1989(83):2372248. [9]SugitaH,OshimaK,TamuraM.Bacterialflorainthegastrointestineoffreshwaterfishintheriver[J].BullJapanSocSciFish,1983,49(9):13872139. [10]尾崎久雄著.李愛杰,沈宗武譯.魚類消化生理[M].上海:科學技術出版社,1985. [11]SakataT,SugitaH,MitsuokaT,etal.Characteristicsofobligateanaerobicbacteriaintheintestinesoffreshwa2terfish[J].BullJapanSocSciFish,1981,47(3):421242. [12]SakataT,SugitaH,MitsuokaT,etal.Isolationanddistributionofobligateanaerobicbacteriafromtheintes2tinesoffreshwaterfish[J].BullJapanSocSciFish,1980,46(10):124921255. [13]RingE,StromTJ.Aintestinalmicrofloraofsalmonids:areview[J].AquacultureResearch,1995(26):7732789. [14]RingE,GatesoupeFJ.Lacticacidbacteriainfish:areview[J].Aquaculture,1998(160):1772203. [15]RingE.Doesdietarylinoleicacidaffectintestinalmi2croflorainArcticcharr,SalíelinusalpinusL.[J].AquacultFishManage,1993a(24):1332135.
| 




































 狗仔卡
狗仔卡


 發表于 2016-12-3 11:24:55
發表于 2016-12-3 11:24:55

 轉播
轉播 淘帖
淘帖 分享
分享 收藏
收藏 支持
支持 反對
反對 提升卡
提升卡 置頂卡
置頂卡 變色卡
變色卡 千斤頂
千斤頂 顯身卡
顯身卡

 發表于 2016-12-14 13:35:33
發表于 2016-12-14 13:35:33






